
Новини
Хижі одноклітинні самостійно винайшли «кулемет Гатлінга»
Навчання
Біологи з університету Британської Колумбії досліджували стрікальний аппатат у клітинах динофлагелят і з'ясували, що його механізм розвивався незалежно від аналогічних апаратів кишечнополісних і влаштований складніше. Деякі динофлагеляти виявилися володарями цілого «кулемета Гатлінга». Стаття опублікована в.
Здатність стрікати жертву або ворога за допомогою спеціальних клітинних пристосувань - відома особливість кишечнополісних тварин, у тому числі різноманітних медуз і поліпів; однак відомі і найпростіші організми, що володіють цією навичкою - динофлагеляти, або панцирні джгутиконосці. Ці рухливі одноклітинні, що мешкають переважно в морських водах, також вміють «вистрілювати» з клітини гарпуноподібними структурами.
Представників планктону часто розглядають як аморфну їжу для великих організмів, проте в реальності між одноклітинними у воді відбуваються цілі війни. Хижі інфузорії, наприклад, за допомогою спеціальних шприців вистрілюють у жертву токсичними речовинами, а їхні жертви обороняються захисними залпами у відповідь, викидаючи на поверхню ніті, що швидко розвертаються, і відштовхуючись від нападника. Разом хижі інфузорії та динофлагеляти поїдають до 60 відсотків первинної продукції водного середовища.
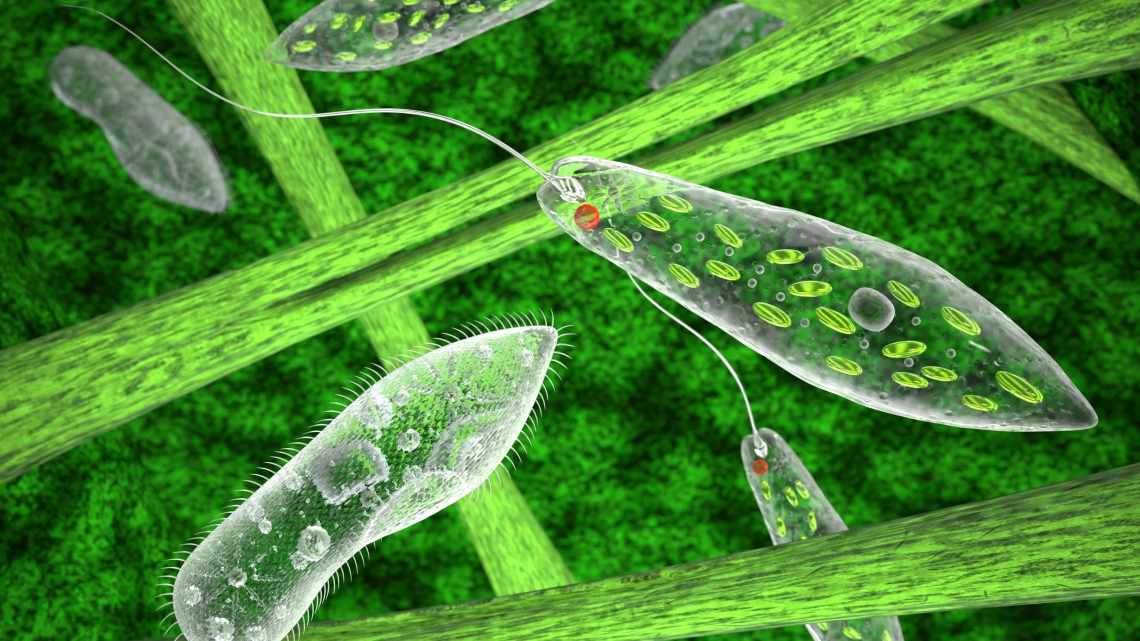
Екструсоми - органели, оточені мембраною і здатні при роздратуванні викидати свій вміст з клітини. Динофлагеляти мають екструсоми декількох типів. Найбільше вважалися схожими на стрікальні апарати кишечнополісних нематоцисти; з їх допомогою полюють на своїх побратимів, наприклад, представники роду. Нематоцисти складаються з капсули, закритої своєрідним затвором, всередині якої лежить закручена в щільний тубус нитка з гострим стилетом на кінці. Нападаючи, клітина викидає такий гарпун у бік жертви, проколює її стилетом і підтягує до себе, щоб поглинути і перетравити.
Раніше вважалося, що механізм дії нематоцист у динофлагелят і кишечнополісних має загальне походження, хоча відповідні структури у одноклітинних у двадцять разів менше розміром. Передбачалося, що кишечнополісні запозичили співвідповідальні гени у динофлагелят - одним з кандидатів на цю роль були представники роду Symbiodinium, які є ендосімбіонтами коралових поліпів.
Вчені шукали гени, схожі з генами стрікального апарату, використовуючи в якості бази даних тридцять геномів і більше ста двадцяти транскриптомів динофлагелят. З'ясувалося, однак, що загальних генів стрікального апарату у динофлагелят і кишечнополісних немає. Зокрема, у динофлагелят відсутні гомологи важливого гена, який кодує осмотичний пропелер полі-лід-глутамат синтазу (відомо, що кишечнополісні придбали його колись шляхом горизонтального транспорту у бактерій). Не виявилося у динофлагелят та інших компонентів стрікального апарату кишечнополісних - мініколлагену, спіналіну та книдоїну, які, мабуть, з'явилися у тварин вже після переходу до багатоклітинної будови тіла. Білки екструсом динофлагелят не продемонстрували схожості ні з якими відомими еукаріотичними білками.
Динофлагеляти, вважають вчені, мають абсолютно унікальний механізм стрікання, який розвивався незалежно від інших подібних систем. За допомогою 3D-реконструкції і даних фокусної іонно-променевої скануючої електронної мікроскопії (FIB-SEM) їм вдалося відтворити структуру цілої клітини, що містить чотири нематоцисти. З'ясувалося, що порівняно зі стрікальними апаратами кишечнополісних вони влаштовані набагато складніше. Якщо у кишечнополісних запираючий механізм капсули влаштований як простий «вилітаючий затор», то у динофлагелят це багатокомпонентна структура, що складається з трьох кілець, і, мабуть, має функції гідравлічної насадки.
Вилітаючи, стилет спочатку проколює цей механізм, проходить крізь насадку, при цьому згорнута в тубулу нитка розправляється і стилет вже зовні проколює тіло жертви. Після цього він розчиняється. Притягує організм жертви до себе, як виявилося, не він сам, а, мабуть, інша нитка, що розташовується в проксимальній частині нематоцисти. Вченим також вдалося поспостерігати дію теніоцистів, інших балістичних структур, які пов'язані структурно з нематоцистами і активуються перед тим, як ті спрацюють. Мабуть, теніоцисти фіксують жертву на місці, щоб нематоцисти могли її проколоти. Координація їх роботи забезпечується спеціальною зв'язуючою органелою.
При подальшому дослідженні динофлагелят з'ясувалося, що незважаючи на загальну базову структуру, механізми стрікання у них можуть різнитися. Представники роду, наприклад, не мають теніоцистів і стиснутих в тубули ниток, замість цього всередині капсули у них розташовуються 11-15 вибудуваних в кільце субкапсул, що працюють як кулемет Гатлінга.
Останнім часом з'являється багато досліджень, що стосуються паралельної еволюції - зовсім недавно ми розповідали, як бактерії набували аналоги ядерних пір незалежно від еукаріот. Вчені вважають, що різноманітність атакуючих і захисних екструсом динофлагелят з'явилася завдяки постійним війнам між хижаками і жертвами, що відбувається в товщі води. У штучних умовах їм вдалося спостерігати, наприклад, як водорость, яка часто є винуватцем токсичного цвітіння води, намагається за допомогою трихосом захиститися від нематоцистів і програє цей бій. Вивчення екструсом може допомогти в боротьбі з подібними організмами при зацвітанні водойм.